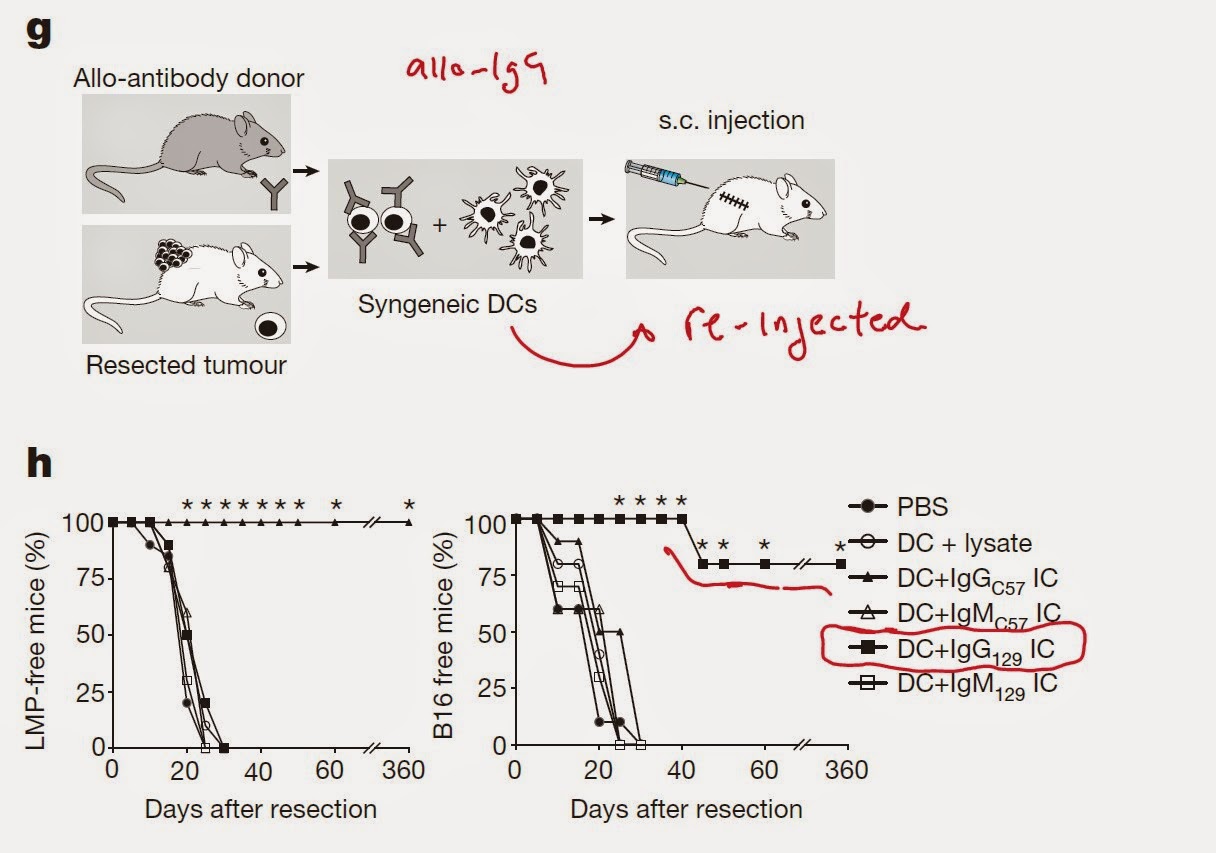
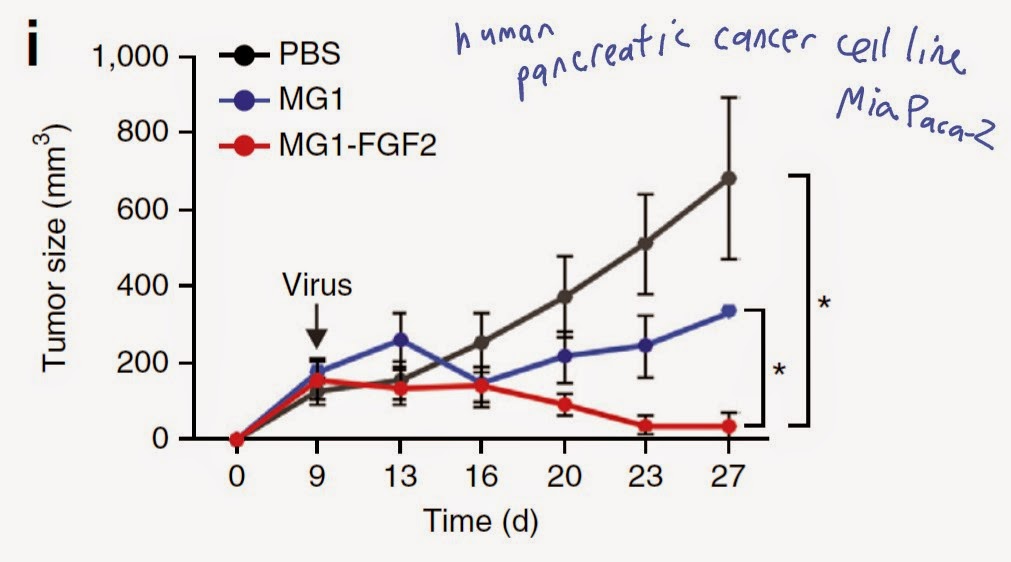
Clonal selection model of immune system ontogeny is the most widely accepted concept of immunology. Simply put, it postulates that T cell clones (B cells too) specific for self-antigens (epitopes) are purged from adaptive receptor repertoire during in utero development.
At the time of its formulation, there was no direct proof for this model. It was mainly supported by observations derived from organ transplantation studies. Later, research on TCR transgenic mice confirmed that self-specific T cells were indeed purged from T cell compartment.
However, more recent studies questioned the validity of results obtain from transgenic mice, mainly due to non-physiological timing of TCR expression in transgenic T cells in the thymus.
In this regard, this new, short study in journal Immunity is of interest. Using sensitive, tetramer-specific T cell enrichment method, the authors showed that human PBMC surprisingly contains equal numbers of both non-self and self-specific CD8 T cell clones.
The authors showed that similar to the frequency of non-specific CD8 T cell clones, healthy human PBMCs contain self-specific CD8 T cell clones with frequency range between 1:104 to 1:106 (see below too).

Next, the authors showed that CD8 T cells derived from both female and male donors contained functionally active male antigen, HY-specific CD8 T cells (though, here CD8 T cells were expanded using non-physiological stimuli, such as anti-CD3/anti-CD28 antibodies or PHA).

However, as expected, further experiments with the pool of enriched self-specific CD8 T cell clones revealed that these self-specific CD8 T cells were anergic when stimulated with specific peptide and anti-CD28 antibody (resembling more physiological stimulation, hopefully).

In summary, these results suggest the following: immune system contains lots of self-specific CD8 T cells. These cells are anergic, i.e. non-responders in an ordinary sense. However, they are capable of becoming functionally active when stimulated by non-conventional ways, like PHA or therapeutic antibodies targeting its signaling molecules (CD28, for example) or during the process of autoimmunity?
Why immune system harbors so many self-specific T cells? It was really surprising that on average, PBMCs from male donors still had 1/3 of number of HY-specific CD8 T cells found in PBMCs from female donors. Even though almost every cell in male donors can express HY antigen, the authors still found so many high-affinity HY-specific CD8 T cells in PBMCs from male donors. Staining for mouse HY-specific CD8 T cells showed the same results.

Another interesting observation was the finding that PBMCs from healthy donors contained very high frequency of pre-proinsulin-specific CD8 T cells (almost 1:104) that were significantly increased in donors with T1D. Is it possible that selection of high frequency of pre-proinsulin-specific CD8 T cells has to due with its cross-reactivity for certain evolutionary relevant pathogen-specific epitope?
David Usharauli

Next, the authors showed that CD8 T cells derived from both female and male donors contained functionally active male antigen, HY-specific CD8 T cells (though, here CD8 T cells were expanded using non-physiological stimuli, such as anti-CD3/anti-CD28 antibodies or PHA).

However, as expected, further experiments with the pool of enriched self-specific CD8 T cell clones revealed that these self-specific CD8 T cells were anergic when stimulated with specific peptide and anti-CD28 antibody (resembling more physiological stimulation, hopefully).

In summary, these results suggest the following: immune system contains lots of self-specific CD8 T cells. These cells are anergic, i.e. non-responders in an ordinary sense. However, they are capable of becoming functionally active when stimulated by non-conventional ways, like PHA or therapeutic antibodies targeting its signaling molecules (CD28, for example) or during the process of autoimmunity?
Why immune system harbors so many self-specific T cells? It was really surprising that on average, PBMCs from male donors still had 1/3 of number of HY-specific CD8 T cells found in PBMCs from female donors. Even though almost every cell in male donors can express HY antigen, the authors still found so many high-affinity HY-specific CD8 T cells in PBMCs from male donors. Staining for mouse HY-specific CD8 T cells showed the same results.

Another interesting observation was the finding that PBMCs from healthy donors contained very high frequency of pre-proinsulin-specific CD8 T cells (almost 1:104) that were significantly increased in donors with T1D. Is it possible that selection of high frequency of pre-proinsulin-specific CD8 T cells has to due with its cross-reactivity for certain evolutionary relevant pathogen-specific epitope?
David Usharauli













.png)
.png)
.png)
.png)